組蛋白乙酰化
1964年,文森特·奧爾弗里及其團隊首次報告了組蛋白乙酰化 [1]。組蛋白乙酰化是一種被廣泛研究的組蛋白后轉錄修飾(PTM),在調控染色質基因轉錄中發揮著核心作用。
1. 什么是組蛋白乙酰化?
組蛋白乙酰化是指將乙酰基添加到組蛋白蛋白質的N-末端賴氨酸殘基上的過程 [1]。所添加的乙酰基的負電荷中和了賴氨酸的正電荷,導致組蛋白與DNA的相互作用減弱,從而使染色質結構更加松弛,有助于轉錄因子接近基因組序列,從而增加基因表達。
已經檢測到各種組蛋白蛋白質上的組蛋白乙酰化。具體而言,H3上的賴氨酸殘基K4、K9、K14、K18、K23、K27、K36和K56顯示出乙酰化,而H4則在K5、K8、K12、K16、K20和K91上顯示出乙酰化。此外,H2A和H2B也經歷賴氨酸乙酰化,在H2A上的K5和K9被修飾,在H2B上的K5、K12、K15、K16、K20和K120被乙酰化。
2. 參與組蛋白乙酰化的酶或蛋白
組蛋白乙酰化是一個動態可逆的過程,由組蛋白乙酰化調節因子(HAMs)協同調節,包括“寫入者”(組蛋白乙酰轉移酶)、“閱讀器”和“擦除者”(組蛋白去乙酰化酶),它們分別負責添加、識別和去除乙酰基。HATs和HDACs之間的這種動態相互作用調節了組蛋白乙酰化的平衡,確保了基因表達模式的精確控制。
2.1 組蛋白乙酰轉移酶(HATs)
HATs,也稱為“寫入者”,負責將乙酰基(-COCH3)從乙酰輔酶轉移到H3或H4的靶賴氨酸殘基的ε-氨基上 [2]。HATS還乙酰化各種非組蛋白底物,因此它們通常被歸類為賴氨酸乙酰轉移酶(KATs)。
基于它們的細胞定位,HAT可分為兩大類:A型HATs位于細胞核,乙酰化核小體組蛋白和其他染色質相關蛋白,而B型HATs位于細胞質,專門乙酰化新合成的游離組蛋白,對轉錄沒有直接影響。
| 組蛋白乙酰轉移酶家族 | 成員 | 定位 | 組蛋白亞基 | 功能 | ||
|---|---|---|---|---|---|---|
| Type A HATs | GNAT | KAT2A (GCN5) | Nucleus | H3K9, H3K14, H3K18, H2B | Transcription activation | |
| KAT2B (PCAF) | Nucleus | H3K9, H3K14, H3K18, H2B | Transcription activation | |||
| MYST | KAT5 (Tip60) | Nucleus | H4K5, H4K8, H4K12, H4K16 | Transcription activation, DNA repair | ||
| KAT6A (MOZ/MYST3) | Nucleus | histone H2B, H3K14 and H4K5, H4K8, H4K12 and H4K16 in vitro and H3K9 in vivo [3] | Transcription activation | |||
| KAT6B (MORF/MYST4) | Nucleus | particularly H3 and H4, and nucleosomal histones with a preference for H4 | Transcription activation | |||
| KAT7 (HBO1/MYST2) | Nucleus | H3K14, H4K5, H4K8, H4K12 | Transcription, DNA replication | |||
| KAT8 (MOF/MYST1) | Nucleus | specifically H4K16 | Chromatin boundaries, dosage compensation, DNA repair | |||
| P300/CBP | KAT3B (P300) | Nucleus | all four histone subtypes: H2AK5, H2B (K5, K12, K15, K20), H3 (K14, K18, K27), and H4 (K8, K12) | Transcription activation | ||
| KAT3A (CBP) | Nucleus | |||||
| Others | Transcription co-activators | KAT4 ( TAF1/TBP) | Nucleus | H3 and H4, H3K14 | Transcription activation | |
| KAT12 (TIFIIIC90) | Nucleus | H3K9, H3K14, H3K18 | Transcription activation | |||
| Steroid receptor co-activators | ATAT1 | Nucleus | - | - | ||
| KAT13A (SRC1) | Nucleus | H3, H4 | Transcription activation | |||
| KAT13B (SCR3/AIB1/ ACTR) | Nucleus | |||||
| KAT13C ( p160) | Nucleus | |||||
| KAT13D (CLOCK) | Nucleus | |||||
| Type B HATS | KAT1 (HAT1) | Cytoplasm | H3, H4, H2A | Histone deposition, DNA repair | ||
| HAT4 (NAA60) | Cytoplasm | H4, H2A | - | |||
表格參考來源:https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4881052/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6311713/
2.2 組蛋白去乙酰化酶(HDACs)
HDACs,也被稱為“擦除者”,是一種能夠從乙酰化的組蛋白中去除乙酰基的酶。組蛋白去乙酰化通過暴露賴氨酸殘基的正電荷,使DNA與組蛋白尾部之間發生相互作用,從而減少了DNA的可及性,導致染色質緊縮和更加致密的染色質結構,有效地抑制了基因表達。
HDACs在各種生理過程中至關重要,包括發育和細胞內穩態,同時也參與了神經退行性疾病、遺傳疾病和癌癥等病理條件的發生。
在人體中,根據它們與酵母的序列同源性,HDAC酶分為四個類別 [4]。I類HDACs包括HDAC1-3和HDAC8。II類HDACs包括HDAC4-7、HDAC9和HDAC10 [5-7]。III類HDACs,也被稱為sirtuins,包括SIRT1-7 [8,9],這些酶在活性時需要NAD+輔酶。IV類HDAC只有一種酶HDAC11 [10]。I、II和IV類HDACs是Zn2+依賴的酰胺水解酶。
| 組蛋白去乙酰化酶家族 | 分類 | 亞族 | 成員 | 細胞定位 | 組蛋白亞基 | 生物學功能 |
|---|---|---|---|---|---|---|
| Classical (Zn2+-dependent) | Class I | HDAC1 | Nucleus | all four core histones [11] | Mediates deacetylation of lysine on the N-terminal part of the core histones and plays and important role in transcriptional regulation, cell cycle progression and development event | |
| HDAC2 | Nucleus | all four core histones [11] | ||||
| HDAC3 | Nucleus | H4K5 H4K12 H2AK5 | ||||
| HDAC8 | Nucleus | preferentially deacetylates histones H3 and H4 [12] | ||||
| Class II | Class IIa | HDAC4 | Nucleus | all four core histones [11] | ||
| HDAC5 | Nucleus | all four core histones [11] | ||||
| HDAC7 | Nucleus | |||||
| HDAC9 | Nucleus | H3K9 H3K14 H3K18 [13] |
||||
| Class IIb | HDAC6 | Cytoplasm | all four core histones [11] | |||
| HDAC10 | Cytoplasm | H3K9 H3K14 H3K18 H3K27 [14] |
Involved in MSH2 deacetylation | |||
| Class IV | HDAC11 | Nucleus | H3K9 H3K14 [15] H4K16 H4K5 H4K12 [16] |
Plays an important role in transcriptional regulation, cell cycle progression and developmental events | ||
| NAD+-dependent | Class III | SIRT1 | Nucleus | H3K9 H3K14 H3K56 H4K16 H1K26 [11] |
Chromatin organization, DNA repair/genome stability, cancer | |
| SIRT2 | Cytoplasm | H4K16 H3K56 [11] |
Chromatin condensation/mitosis, DNA repair, cancer | |||
| SIRT3 | Mitochondria | H4K16 [11] | Chromatin silencing, DNA repair, cellular stress | |||
| SIRT4 | Mitochondria | / | Regulates the cellular metabolic response to DNA damage | |||
| SIRT5 | Mitochondria | / | Regulates the mitochondrial lysine succinylome and metabolic networks | |||
| SIRT6 | Nucleus | H3K9 H3K56 [11] |
Telomeric chromatin/senescence, DNA repair/genome stability, energy metabolism | |||
| SIRT7 | Nucleolus | H3K18 [11] | Mediates deacetylation of H3K18ac Mediates deacetylation of lysine residues on the N-terminal part of the core histones |
表格參考來源:https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3970420/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7444376/
2.3 讀取蛋白
組蛋白乙酰化讀取蛋白特異地識別乙酰化的賴氨酸,并招募轉錄因子和其他調節蛋白與DNA結合,啟動轉錄,從而激活基因表達 [17,18]。它們通常是含有溴結構域(BRD)的蛋白或者是乙酰化賴氨酸結合蛋白,比如BRD和額外末端結構域(BET)家族,負責將乙酰化信號解釋為基因表達和染色質重塑中的特定變化。
BRDs突出顯示為唯一能夠識別和結合乙酰化組蛋白賴氨酸殘基的蛋白群。它們廣泛分布在各種組織中,并根據序列或結構相似性被分為八個家族,每個家族都顯示出多樣的活性,比如組蛋白修飾和染色質重塑。
| BRDs | Members |
|---|---|
| Class I | BAZ1A, BPTF, CECR2, GCN5L2, PCAF |
| Class II | BRD2, BRD3, BRD4, BRDT |
| Class III | BAZ1B, BRD8B, BRWD3, CREBBP, EP300, PHIP, WDR9 |
| Class IV | ATAD2, ATAD2B, BRD1, BRD7, BRD9, BRPF1, BRPF3 |
| Class V | BAZ2A, BAZ2B, SP100, SP110, SP140, SP140L, TRIM24, TRIM33, TRIM66 |
| Class VI | MLL, TRIM28 |
| Class VII | BRWD3, PHIP, TAF1, TAF1L, WDR9, ZMYND8, ZMYND11 |
| Class VIII | ASH1L, PBRM1, SMARCA2, SMARCA4 |
表格參考來源:https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7686570/
3. 組蛋白乙酰化的機制
組蛋白酰轉移酶(HATs)酰化組蛋白賴氨酸殘基,減少了組蛋白和DNA之間的靜電相互作用,導致染色質結構松弛。因此,DNA被暴露出來,有助于轉錄激活因子(TA)被招募到基因組序列上,激活基因表達。
相反,組蛋白去乙酰化酶(HDACs)可以與轉錄抑制因子(TR)復合物相互作用,去除這些乙酰修飾。這增強了DNA和組蛋白之間的靜電相互作用,導致緊湊的染色質,抑制轉錄。保持組蛋白乙酰化和去乙酰化之間的平衡對于正確的基因表達非常關鍵。
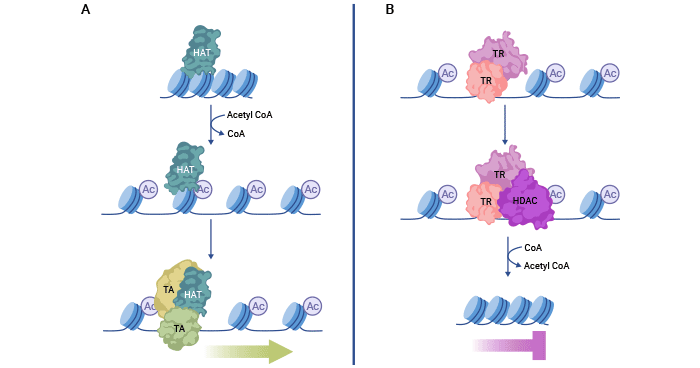
圖1. 組蛋白乙酰化與去乙酰化機制
圖片參考來源:https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7820808/
4. 組蛋白乙酰化的功能
大量研究數據表明,組蛋白乙酰化在細胞中廣泛存在,并參與各種細胞活動,包括核小體組裝的調節、染色質的折疊和解構、基因轉錄、DNA損傷修復、細胞周期和凋亡 [19]。
5. 與其他修飾的相互作用
組蛋白乙酰化與其他組蛋白修飾之間存在復雜的相互作用,比如甲基化、磷酸化和泛素化。它們可以相互合作或拮抗,影響染色質結構和基因表達模式。
在釀酒酵母薩克羅米瑟斯小麥,Snf1激酶介導的H3S10ph促進了由Gcn5乙酰轉移酶介導的H3K14的乙酰化,在基因激活期間增強了組蛋白H3與14-3-3蛋白Bmh1和Bmh2的相互作用 [20]。在哺乳動物細胞中,H3K18ac和H3K23ac促使CARM1甲基轉移酶介導的H3R17甲基化,導致雌激素響應基因的激活 [21]。
組蛋白修飾的相互作用也可以在去除特定修飾方面發揮作用。例如,在釀酒酵母中,與RNA聚合酶II結合的Set2甲基轉移酶甲基化H3K36,引導核小體在RNA聚合酶通過后由Rpd3S脫乙酰酶復合物去乙酰化H3和H4 [22]。
Wojcik等人證明,H2A的多個位點,包括K5、K9、K13和K15的乙酰化抑制了H2BK120的泛素化 [23]。
6. 發育和疾病
組蛋白乙酰化參與了許多生物學過程,包括發育、分化和對環境信號的響應。組蛋白乙酰化的失調與各種疾病相關,比如癌癥、神經系統障礙和代謝性疾病,突顯了它在疾病發病機制中的重要性。
在癌細胞中觀察到異常的組蛋白乙酰化模式,導致細胞不受控制地增長和腫瘤發生。HDAC抑制劑能夠恢復適當的乙酰化模式,被認為是潛在的治療某些癌癥的方法,提供了一種新的方式來靶向癌細胞而不傷害正常細胞。
在神經退行性疾病中,比如阿爾茨海默病和帕金森病,組蛋白乙酰化的紊亂與與神經細胞存活和功能相關的基因表達模式的變化有關。失調的組蛋白乙酰化可能導致神經退行性病變和疾病進展。
此外,組蛋白乙酰化還與代謝性疾病相關,它可能影響與葡萄糖代謝和脂質穩態有關的基因的表達。對飲食和環境因素的反應引發的表觀遺傳變化可能導致代謝失調和與肥胖相關的疾病。
總的來說,HATs、HDACs和BRDs形成了一個復雜的調節網絡,控制組蛋白的乙酰化狀態,從而影響基因對轉錄的可及性。這種表觀遺傳機制在各種生物學過程中發揮著關鍵作用,包括發育、分化和對環境信號的響應。這些酶的失調可能導致基因表達模式的異常,與多種疾病相關,使它們成為治療干預和藥物開發的有希望的靶點。
參考文獻:
[1] Allfrey VG, Faulkner R, Mirsky AE. Acetylation and methylation of histones and their possible role in the regulation of RNA synthesis [J]. Proc Natl Acad Sci USA. 64;51:786–794.
[2] Roth, S.Y.; Denu, J.M.; Allis, C.D. Histone acetyltransferases [J]. Annu. Rev. Biochem. 2001, 70, 81–120.
[3] Perez-Campo FM, Costa G, et al. The MYSTerious MOZ, a histone acetyltransferase with a key role in haematopoiesis. Immunology. 2013 Jun;139(2):161-5.
[4] Seto E, Yoshida M. Erasers of histone acetylation: the histone deacetylase enzymes [J]. Cold Spring Harb Perspect Biol. 2014;6(4):a018713–a.
[5] Grozinger CM, Hassig CA, Schreiber SL 1999. Three proteins define a class of human histone deacetylases related to yeast Hda1p [J]. Proc Natl Acad Sci 96: 4868–4873.
[6] Kao HY, Downes M, Ordentlich P, Evans RM 2000. Isolation of a novel histone deacetylase reveals that class I and class II deacetylases promote SMRT-mediated repression [J]. Genes Dev 14: 55–66.
[7] Kao HY, Lee CH, Komarov A, Han CC, Evans RM 2002. Isolation and characterization of mammalian HDAC10, a novel histone deacetylase [J]. J Biol Chem 277: 187–193.
[8] Frye RA 1999. Characterization of five human cDNAs with homology to the yeast SIR2 gene: Sir2-like proteins (sirtuins) metabolize NAD and may have protein ADP-ribosyltransferase activity [J]. Biochem Biophys Res Commun 260: 273–279.
[9] Du J, Zhou Y, et al. 2011. Sirt5 is a NAD-dependent protein lysine demalonylase and desuccinylase [J]. Science 334: 806–809.
[10] Gao L, Cueto MA, Asselbergs F, Atadja P 2002. Cloning and functional characterization of HDAC11, a novel member of the human histone deacetylase family [J]. J Biol Chem 277: 25748–25755.
[11] Seto E, Yoshida M. Erasers of histone acetylation: the histone deacetylase enzymes [J]. Cold Spring Harb Perspect Biol. 2014 Apr 1;6(4):a018713.
[12] Hu E, Chen Z, et al. 2000. Cloning and characterization of a novel human class I histone deacetylase that functions as a transcription repressor [J]. J Biol Chem 275: 15254–15264.
[13] Kailin Yan, Qiang Cao, et al.Histone Deacetylase 9 Deficiency Protects against Effector T Cell-mediated Systemic Autoimmunity [J]. Immunology, Volume 286, Issue 33, 19 August 2011, Pages 28833-28843.
[14] Li Y, Peng L, Seto E. Histone Deacetylase 10 Regulates the Cell Cycle G2/M Phase Transition via a Novel Let-7-HMGA2-Cyclin A2 Pathway [J].







